Collecting tips
It is often difficult to separate Hebeloma in the field because so many are macroscopically similar. “Hebeloma hotspots” are a reality. If in any doubt it is preferable to make separate collections to avoid the many mixed collections that we regularly receive and have often collected ourselves. In the field, we carry a notebook, collecting box with each compartment pre-labelled, GPS, camera, x10 hand lens and penknife. We take a photograph in situ, record the habitat and potential mycorrhizal partners, the number of basidiomes in the collection and any characteristics that might disappear or change during handling: e.g. smell, drops on the lamellae, floccosity of the stipe. We record the GPS location including the altitude. It is also useful to record the colour of the pileus during daylight hours. Young basidiomes can look quite different from their mature state and tend to be uniform in colour but more mature specimens may exhibit two or more colours, so these are also noted. We use Kornerup & Wanscher (1961) as our colour charts and equate swatch numbers with terms introduced by Vesterholt (2005).
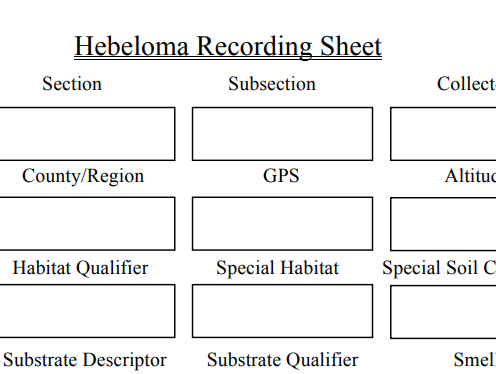
The recording sheet allows systematic
collection of data in the field
We transfer the field information to a recording sheet with fields that match those in our database. Each collection is given a unique identifier (ID) which corresponds to the entry in our database and all associated material such as photographs, spore prints, DNA sample and sequences. The recording sheet is divided into sections. The first covers the collection site and includes: field determination, collectors name, ID, place collected, country, GPS position, altitude, date of collection, habitat data including associated organisms, smell and taste (rarely recorded, given toxicity concerns, and not used in our keys). Next we examine and measure the pileus and record: shape (applanate, convex, umbonate (weakly, broadly, strongly) or umbilicate) and whether any regular spotting occurs on the pileus, whether any veil remnants exist, whether it is rugulose or pruinose, if the cap is hygrophanous, colour and any features on the pileus margin (eroded, striate, crenulate, overhanging the lamellae, scalloped, smooth, wavy. The attachment of the lamellae does not appear to be particularly important, but we do record it (emarginate, adnexed or adnate, rarely decurrent) however drops on the lamellae are a good character. We note if they are absent, visible to the naked eye or only with a hand lens (indicating type of cheilocystidia). A white fimbriate lamella edge is significant, and the number of full-length lamellae is a very important character. We tend to view under a large magnifying glass, count a quarter of the lamellae then multiply by 4 and we do this for each basidiome.
The shape of the stipe is recorded (bulbous, subbulbous, clavatae, cylindrical or tapering (referring to the shape towards the base). The stipitipellis is recorded (fibrillose, velutinous, pruinose, pruinose at apex, floccose, floccose at apex). The stipe of one good basidiome should be cut to expose the interior and recorded as hollow (often when mature) or stuffed and the presence of a basal or superior hanging wick. Discolouration sometimes occurs in the lower part of the stipe in situ or with handling and is worth noting. The base may show signs of rooting or a conspicuous number of thick rhizoids, so these characters should be recorded.
Measurements are made, ideally with callipers and a magnifying glass for accuracy. The following data are recorded for all mature species: pileus width, stipe length, width in the stipe median and stipe width at base. Any additional information or sketches are made on the reverse of the recording sheet and entered into the database within a free-format notes field.
A spore print is made using the cut basidiome. We use the same basidiome for microscopic and molecular work to ensure that all data is coming from the same source. For the spore print we place the cut sections between two microscope slides placed across the top of a small pot or jar (for example a yogurt pot). A second pot is placed on top of the slides to retain moisture and left overnight (if possible). The following morning, the slides with the spore deposit are put into a slide carrier. In the lab the spores from one slide are scraped across the slide leaving sufficient spores for microscopic analysis. The remaining spores are added to the second slide and scraped into a central pile. A cover slip is placed over the spore pile and taped down and then used to determine the spore colour.
Before drying the basidiomes, a small fragment of pileus and lamellae is broken off and placed in a vial containing CTAB (Largent & Baroni 1988) for potential DNA analyses. All basidiomes are then dried using a food dessicator at a temperature of 40-45 degrees C for at least 5-6 hours (typically overnight). This temperature is selected to preserve the DNA; they need to dry fast enough that they do not rot, but not so that they cook! It is important to label each collection as it is placed in the dryer. Dried basidiomes are placed in zip locked bags and labelled with their database ID along with all supporting materials (recording sheet, sample vials etc.).
a. Macroscopic characters
Basidiome size:
Size is a very important character. Pileus diameter (p) can range from very small (7 mm) to large (up to 150 mm). Stipe width (w) measured at its median, and stipe length (l) should also be recorded. We can then calculate stipe Q (the ratio of length and width i.e. l/w) and slenderness (s) which is the square of the length of the stipe divided by the product of the pileus width and stipe width (i.e. (l x l) /p x w)). Always use mature specimens for calculating (s). A slim basidiome will have a high slenderness value and a short, squat basidiome a much lower value. For example, Hebeloma pusillum (s) values range from 12 to 65 whereas H. sinapizans has a maximum value of 14 and H. erumpens' highest value is 4.4.
Pileus:
The pileus shape is hemispherical and expanded in young specimens. The presence of a broad or narrow umbo is characteristic of many species although the pileus can become umbilicate with age. The involute margin seen in some young species rarely remains, but margins may be crenulate (often in H. birrus and H. cavipes) upturned, wavy or eroded and sometimes the pileus overhangs the lamellae. The pileus surface is mainly smooth (although sometimes pubescent looking with lingering remains of a universal veil), with only H. pubescens where the pileus is covered in hairs and section Sacchariolentia, in particular H. odoratissimum, being tomentose to squamulose. Hebeloma have a distinct ixocutis causing a viscid to slimy pileus but this can dry out quickly but leaving leaves and other debris stuck on the pileus. A grey, uneven pruinose layer is often found in H. sects. Naviculospora and Theobromina. The colour of the mature pileus varies from white to grey and brown with occasional yellow, orange, pink or red tones. Some species have a pileus more or less uniformly coloured while others have a distinct two-three coloured pileus, more or less in zones. For taxonomic purposes the colour at the centre of the pileus is of most importance. Young basidiomes tend to have a more uniform appearance and the colour is often more intense. Spotting, usually close to the margin, is not uncommon.
Lamellae:
The attachment of the lamellae has not, at least in European species, been found (taxonomically) to be of importance and neither has the depth (although this is a reasonably consistent character within a species) but we always record these characters. Lamellae tend to be emarginate but may be adnexed, adnate or (rarely) decurrent. The number of full-length lamellae (L) is, however, a very important character. Young species can have white or greyish to brownish lamellae but mature specimens are usually dull brown in colour with the exception of members of section Porphyrospora, e.g. H. porphyrosporum, which have a reddish tinge. A white fimbriate edge on the lamellae, a sterile edge, is usually indicative of clavate to capitate cheilocystidia whereas this white edge is missing or less distinct when the cystidia are less differentiated from the basidia or have a small apex. White lamellae are rare but do occur in sterile basidiomes or where the spores are not fully formed and were formerly described as a separate genus: Hebelomina.
Droplets of varying sizes may form on the edge of lamellae in some species and are a useful character that generally reflects the shape and apical size of the cheilocystidia. The absence of droplets is also useful; so, for example, a large Hebeloma (L > 100) found in Europe without drops is likely to be H. sinapizans.
Stipe:
Most species are cylindrical to clavate in stipe shape, but some typically have a bulbous base, particularly common in H. sects. Sinapizantia and Velutipes. Some have an ability to be strongly rooting while others only slightly so or are fusiform-pointed. In such cases, the base of the stipe often tends towards a turnip shape, widening at the end before tapering off. These rooting structures have, in many cases, been linked to subterranean mammalian nests or carcasses - see e.g. Sagara (1980) and Sagara et al. (1981). While the nature of the stipe base is very important the character is not always consistent.
The stipe is always dry and slightly to coarsely fibrillose. The presence of caulocystidia is a very useful character and make the stipe appear velutinous, pruinose and floccose. For example, the stipes of H. crustuliniforme and H. sinapizans are highly floccose and may look quite different from the velutinate stipes of, say, H. leucosarx and H. velutipes. Stipe colour may change as species mature and, in some cases, browning develops from the base upwards (Smith et al. 1983). The interior of young basidiomes is usually stuffed but tends to become hollow with age. One exception is found in H. alpinum which remains stuffed. A basal or superior hanging wick is present in some species.
Flesh:
The colour of the flesh is usually white, yellowish or some shade of brown. The stipe flesh may discolour as it matures.
Presence of a veil:
A visible veil is an important taxonomic character. It appears that all Hebeloma start out with a cobweb-like to woolly or membranous universal veil, usually only seen in primordia. But it can leave traces on the cap and the lower part of the stipe. For example, in H. radicosum it leaves a thick and membranous ring on the stipe; in H. pumilum it leaves more cortina-like remnants on the stipe.
Sometimes, a cobweb-like cortina between the pileus margin and stipe is clearly visible.
This feature is usual in H. sect. Hebeloma and often present in H. sect. Scabrispora.
Taste and smell:
Most Hebeloma (H. sects. Denudata, Hebeloma, Sinapizantia, Theobromina, Velutipes) have a raphanoidradish-like smell but a basidiome may need to be cut before it becomes apparent. Conversely, H. sect. Scabrispora offers a range of scents from tea, cocoa and chocolate to soap while H. sect. Myxocybe is described as having a marzipan-like smell. The strong, sweet, fragrant smell of H. sect. Sacchariolentia is distinctive and consistent and therefore a very a good character.
While taste may be a useful taxonomic character, owing to the suspected toxicity of many species of Hebeloma, this character is rarely recorded and not used taxonomically.
Spore deposit:
The spore colour, in mass, varies from umber to dark brick. Exceptions, with a reddish brown deposit, are found in members of H. sect. Porphyrospora.
Exsiccata characters:
The exsiccate can become blackened and hard. This feature can be affected by the wetness of the basidiome when collected or by the drying facilities but is often seen in H. sordescens and some species of H. sect. Sacchariolentia. Most species that exhibit brown colouring when living tend to remain brown.
b. Microscopic characters
All photographs and measurements presented here are based on the microscopical analysis of dried material. We use a Leica DMRZA2 microscope system with a Leica DFC495 camera connected to a computer running Leica Application Suite (LAS) V4 image capture and image analysis software. All relevant characters are photographed, measured and analysed using the image analysis software and automatically fed into spreadsheets where statistics are calculated and transferred to the database. This means that all measurements are stored and can be repeated. Photographs to be used for measurements are taken at x500 or x1000 magnification. Important characters such as basidia, caulocystidia, cheilocystidia are all photographed at x1000 and spores are photographed at x1600 with a focus on both the centre, to illustrate the shape and the top to show ornamentation.Spores:
Dimension and shape: Shape ranges from ellipsoid to cylindrical, ovoid, amygdaloid, limoniform or fusoid and even navicular. Distinct papilla at the end opposite the hilar appendage are absent for some species but appear weakly or strongly in other species. Spore colour is worth noting if very pale, yellow-brown or distinctly brown. Spore size ranges from 6.5 x 4 µm (small) to 16 x 9 µm (large). The spores are usually rough, to a greater or lesser extent, with ornamentation within the epitunica (perispore).
The spore classification used here is based on ideas proposed by Vesterholt (2005), Boekhout (1982) and others, which focus on ornamentation, loosening perispore and dextrinoidity.
Spore character measurements:
Firstly, every microscope is different, so it is very important to ‘self-calibrate’ with your own equipment on known material, to understand these measures better.
Ornamentation:
This is a consistent character for all ripe spores and can be classified as follows:
- O1: spores almost smooth, even under immersion
- O2: spores very weakly ornamented, only visible under immersion
- O3: spores distinctly ornamented, ornament visible without immersion but not conspicuous
- O4: spores with fairly strong ornamentation, always easy to see without immersion
Loosening of the perispore:
The loosening of the perispore (which is indextrinoid) appears as a hyaline sack or blisters under a light microscope and can be seen clearly in Melzer’s reagent. It is highly variable and depends on maturity. We have developed a scale:
- P0: perispore not loosening
- P1: perispore somewhat loosening (in a few to all spores), immersion often needed
- P2: perispore distinctly loosening (in few to all spores), also visible without immersion
- P3: perispore strongly and consistently loosening. Species with strong spore ornamentation often, but not always, also exhibit a loosening perispore
Dextrinoid reaction:
A dextrinoid reaction with Melzer's reagent is a very important character, but intermediate degrees of reactions often occur, so we have developed a five-point scale:
- D0: spores completely indextrinoid
- D1: spores with an indistinct brownish tint
- D2: spores weakly but distinctly dextrinoid, becoming pale brown or yellow brown
- D3: spores rather strongly dextrinoid, becoming medium brown
- D4: spores strongly dextrinoid, immediately becoming deep and intensely red-brown.
Photographing and measuring spores:
Ideally spores are photographed and measured from a spore print, but where unavailable, from a fragment of exsiccata lamella or from the top of the stipe. The dried material is mounted in Melzer’s reagent. A few moments are allowed for any colour change to settle. Enough photographs are taken at x500 to measure 50 spores (usually 2-3 photographs); more images are needed if photographing from the lamellae or the top of the stipe (5-12).
The image analysis software is set up to detect and trace, automatically, the shape of the spores for measurement and so, for example, will exclude any that are not focused for the entire length of the spore. Among the measurements recorded are: length (l), width (w), Q and area. Spore statistics are automatically computed to provide 5% and 95% percentiles, mean, median and standard deviations for each parameter. These data are automatically transferred to the database. More photographs are taken of spores mounted in 5% KOH at x500 and the colour recorded: hyaline, pale, pale brown, yellow, yellow brown or brown. Photographs are also taken at x1600 with spores mounted in both Melzer’s and 5% KOH thus providing high definition photographs of the spore shape, the ornamentation of the spores and other characters such as any loosening perispore.
During this process the spores are classified as: ellipsoid, cylindrical, ovoid, amygdaloid, limoniform, fusoid or navicular, the OPD values of the spores are estimated, whether a papilla is present and whether the spores are guttulate.
If measuring the spores manually, it is still recommended that at least 30 spores are measured in order to generate reasonable statistics.
Basidia:
Basidia are mostly subclavate to subcylindrical, usually with four sterigmata and with or without a weak constriction in the middle. Occasionally basidia with one, two and three sterigmata are found. The basidia are photographed at x500 and/or x1000. Length and width near the apex are measured and the Q value (l/w) is determined; averages for these parameters are then calculated. Basidia are not used in our traditional keys or identifier but they can be a useful character, providing further evidence of a determination, (e.g. H. lutense has very long, narrow basidia).
Cystidia:
Cheilocystidia:
The shape of the cheilocystidia is a very important character. Many Hebeloma have crowded cheilocystidia and a sterile lamella edge, but there are exceptions such as within H. sect. Scabrispora where they can be sparse and located on the lamella edge, together with the basidia. It is important to consider a number of cystidia to determine the dominant shape. Five primary shapes are defined: cylindrical; ventricose (including lageniform); clavate near the apex and gently tapering downwards; clavate-stipitate (usually clearly constricted below the apex, sometimes with the base widening slightly near the very bottom); clavate-lageniform (often looking almost hourglass, widened near the apex, constricted in the central part and widened again in the lower half). In the last two cases, some cystidia may be verging on capitate or spathulate rather than clavate. Cheilocystidia tend to be thin walled and hyaline but sometimes there is some thickening of the walls and this can be a useful character (e.g. H. helodes exhibits apical thickening). To determine the shape the following measurements should be taken: length (l), maximum width near the apex (A), minimum width in upper part, below the apex (M), maximum width in lower third (B), written as l x A x M x B and the following ratios calculated: A/M, A/B, B/M.
To examine and measure this important character cheilocystidia are mounted in either Melzer’s reagent or 5% KOH (the latter usually allows easier separation and hence measurement). The dominant shape is recorded as well as any other special features that occur more than once, for example, septate, geniculate, sinuate, bifurcate, wall thickenings. In some cases the cheilocystidia appear quite irregular and it is necessary to examine a large number before forming an opinion on which shape is the dominant one. At least 20 cheilocystidia are measured, ideally separated from the lamella edge so that the features are clear. These are measured individually and averages are calculated and transferred to the database. A very useful character for species determination, particularly in sections such as H. sects. Denudata and Velutipes, is the average width of the cheilocystidium apex, A. Ideally, some 100 are measured on the lamella edge while trying not to be selective.
Where the lamella edge is sterile the cheilocystidia are usually crowded along the edge of the lamellae. To measure sufficient cystidia to be able to calculate a reliable average for the value A typically requires 5-6 photographs at x500 magnification, or more at x1000. Where the lamella edge is not sterile, a few extra photographs may be necessary. To measure l, A, M and B for a cheilocystidium, the complete cheilocystidium must be within view and this may mean gently tapping the slide to release a few from the lamella edge. These are usually photographed at x1000 to see clearly the shape and any special features. A few Hebeloma have more than one type of cheilocystidia (e.g. H. sect. Velutipes). In such cases, as far as possible, a representative sample for measurement is selected. It is sometimes difficult to separate the cheilocystidia from the basidioles (H. sects. Scabrispora or Sacchariolentia); in such cases an examination of the caulocystidia first, which may be more easily distinguished, allows familiarization with the cystidium shape before attempting to measure the cheilocystidia.
Pleurocystidia:
Pleurocystidia are not common in Hebeloma but they do occasionally occur (e.g. H. cistophilum and H. sinapizans and they are reasonably common within H. sect. Porphyrospora). They are often similar to the cheilocystidia and hence may easily be confused with cheilocystidia. To avoid this, when searching for pleurocystidia, it is safest to take a segment of the lamella and cut the edge away before searching for them.
Caulocystidia:
All Hebeloma have caulocystidia. For some species they are limited to the zone between the cortina and the top of the stipe (e.g. H. sect. Hebeloma), while for others they are found the length of the stipe. They often tend to appear in bundles giving the stipe a floccose or pruinose appearance. They are usually similar to the cheilocystidia but more irregular, larger and often multi-septate. We photograph them at x500 and x1000, mounted in 5% KOH. The caulocystidia have not been used as a taxonomic character in the keys to Hebeloma species.
Pellis structure:
An ixocutis is formed in the upper layer of the pileus. The epicutis comprises narrow, hyaline to yellowish, smooth to encrusted hyphae embedded in a gelatinous layer. The hyphae may be erect (ixotrichoderm) but are often indistinguishable from the surrounding matrix particularly in dried material. The depth of the epicutis ranges from less than 30 µm in almost dry species to 250 µm in viscid species such as H. mesophaeum and can be a very useful taxonomic character. The subcutis is formed by nearly hyaline to dark brown elements often with a resinous parietal pigmentation.
This nature and size of the ixocutis is an important character. To prepare a slide, a few very thin vertical sections are cut through the top of the pileus of the dried specimen and the thinnest are examined, first, without a cover slip, in a drop of 5% KOH, either under a binocular microscope, at x80 or under the compound microscope at x125, and photographed. A cover slip is then added and the magnification increased to x500 to observe the hyphae and measure their width. The presence or absence of encrusted hyphae, the colour and shape of the elements that make up the subcutis and the trama immediately below the subcutis are all noted, photographed and measured as appropriate.
Trama:
The trama below the cutis is comprised of cylindric to ellipsoid or sausage-shaped elements of varying thickness and hyphae colour can range from hyaline to distinctly yellow or brownish. The brown pigmentation can be very intense.
The trama of the hymenium (examined in 5% KOH), may also be hyaline, pale brown or brown. In our experience this character is very subjective and dependent on the thickness of the preparation.
Clamp connections:
Clamp connections have been found in all collections.